


摘 要:鉴于因果建模领域突出进展,因果模型可为先天/后天之争的解决提供一个形式化的语义系统。
关键词:中国社会科学;中国社会科学杂志社;中国社会科学评价;先天/后天之争;发育系统论;遗传分析;因果模型

摘要:达尔文的自然选择为生命的目的提供了机械论解释,是当代生物学最为突出的因果特征。然而,这一特征却与发育视角下把基因和环境作为表型的共同原因的主张,貌似产生了冲突。先天/后天之争在当代生物学的持续讨论,揭示了基因在发育和演化两个维度的因果张力。梳理此争论的不同呈现形式及其发展,可以看到,更为细致的因果相关概念被逐渐澄清,但仍缺少一个基础的因果语义框架。鉴于因果建模领域的突出进展,因果模型可以为先天/后天之争的解决提供一个形式化的语义系统。此外,因果模型及其计算可以从形式上说明因果认知的规范性面向,因而有望为生命之目的给出一种彻底的自然主义诠释。
关键词:先天/后天之争 发育系统论 遗传分析 因果模型
作者陆俏颖,北京大学哲学系助理教授(北京100871)。
一、目的、功能与结构
现代生物学是研究生命的科学学科,目前形成了许多分支。从生命的组成层级来看,分子生物学和生物化学研究生命分子的化学基础,细胞生物学、胚胎学和发育生物学研究更高层级的组织形态,生态学、古生物学和系统发育学则将生命体放置于更为广阔的时空尺度。每个分支学科都有各自的专业词汇、研究问题与研究框架。鉴于此,我们如何可以普遍地谈论生物学中的因果问题呢?事实上,由于因果概念在遗传和演化上的特殊性,因果问题在生物学中尤为突出,是生物学哲学的传统问题,也是近期讨论的热点。其中的缘由还需从目的论说起。
我们对于生命的理解,与非生命对象相比,总是绕不开目的论。除了复杂生物所具有的以目标为导向的行动能力(例如,因为饥饿所以捕食),生命本身似乎就蕴含了内在的目的。常言道:“种瓜得瓜,种豆得豆。”生命的传承意味着内在目的的传递,传承而来的目的又决定了生命个体的发育轨迹,使之产生与周围环境相契合的表型性状。在现代科学的机械观看来,生命所具有的目的及其与环境的适应性似乎无从解释。康德在《判断力批判》中这样说道:“……也许有朝一日还会出现一个牛顿,他按照不是任何意图所安排的自然法则来使哪怕只是一根草茎的产生可以理解,这对于人类来说也是荒谬的……” 然而,如此“荒谬的”事情达尔文却做到了:一根草茎产生的背后没有真正的目的,有的是基于自然选择的演化过程。
一般认为,自然选择的发生需要具备三个条件:(1)种群中不同个体之间存在性状上的差异(即表型变异),如某动物群体中同时存在具有和不具有类似于心脏器官的个体;(2)给定种群所处的环境,性状差异导致个体具有不同的适应度(即差别性适应度),如具有类心脏器官的个体泵血能力更强,因而更容易觅得食物,留下更多的后代;(3)具有不同适应度的个体,其性状可以遗传(即适应度可遗传),如具有类心脏的个体更有可能生育具有类心脏的后代。若上述三个条件被满足,一种选择过程会自然地发生:在下一代群体中,拥有高适应度的个体(即具有类心脏器官的个体)的占比就会上升;经过长期累积,最终所有个体都会具有类心脏器官,因而都具备泵血的能力。
在上述演化解释中,心脏因其能泵血而被自然选择所青睐,继而被保留下来。这解释了现在心脏为何存在。虽然泵血和嘣嘣声都是心脏存在而导致的效应,但只有前者才是心脏被选择的真正原因。心脏所具备的泵血功能,使得心脏的存在具有了某种目的性,即心脏存在是“为了”泵血。在这个意义上,生物的内在目的被还原到了性状的功能上。这开启了生物学哲学中关于功能概念的讨论, 但其理论后果远远不止于此。与具体的物理结构相比,功能才是自然选择的直接目标。原则上,无论物理结构的差异有多大,只要能够实现相似的功能,那么自然选择就会一视同仁地对待。因而,自然选择对于结构是“盲目的”。达尔文演化论作为现代生物学的范式理论,使得生物学中的重要概念几乎都通过功能来定义。例如,翅膀、鳍等,甚至基因的概念也是如此。由于某个特定功能原则上可以由无限种类的结构来实现,一个功能无法被还原或被等同于某个物理结构的集合。这从根本上导致了生物学中结构与功能的理论割裂。
20世纪 40 年代,达尔文演化论与孟德尔遗传学的结合催生了“现代演化综合”(Modern Synthesis)。现代综合的一个重要理论预设是,对于演化或系统发生学(phylogeny)的研究不需要指向发育或个体发生学(ontogeny)。此预设后因迈尔(E. Mayr)对于近因和远因的区分而被广泛接受。迈尔认为,生物学的问题可以分为两大类。一类是“为何”问题,如蝙蝠为何具有翅膀?这类问题询问的是远因,即性状功能的演化历程,其中自然选择是最重要的远因;另一类是“如何”问题,如蝙蝠的翅膀如何在发育中产生?该问题询问的是近因,即性状结构的产生机制。两类问题的区分造就了两个领域的区分:演化论者研究演化以回答“为何”问题,分子生物学家研究发育以回答“如何”问题。可见,功能与结构的理论割裂使发育从演化图景中被割舍,但也因此埋下了因果推断的隐患。
20世纪80年代兴起的演化发育生物学(evo-devo),研究个体发育与群体表型变化之间的关系,尤其是发育如何为演化提供了表型变异。该学科曾被寄希望于颠覆演化理论对于发育的忽视,但最终并未成功。随着生物学分支领域的发展,尤其是表观遗传学、生态学、发育生物学,越来越多的经验证据指向了发育对于演化的重要影响。近因和远因的二分框架逐渐成为探索发育与演化、结构与功能之交互问题的理论瓶颈。回顾类心脏的演化案例,发育过程产生了新的类心脏结构,自然选择青睐于泵血功能,使具有类心脏的个体得以繁衍,繁衍的后代又在发育过程中产生了类心脏。这里,遗传是连接发育和演化的真正桥梁。因此,可遗传的基因及其对于表型的因果角色成为了争论的焦点。
本文以生物学哲学的视角,从基因的因果性问题入手,梳理生物学中因果相关理论的变迁及其未来的发展趋势。本文第二节聚焦于先天/后天之争的理论面向及其发展,揭示基因在发育和演化两个维度的因果张力。第三节介绍此争论的科学方法论面向及其进展。随着讨论越来越具体化,与生物学尤其相关的因果概念被逐渐澄清。虽然概念分析能够帮助我们进一步理解基因的因果张力,却无法澄清或消解这一张力。本文第四节借鉴因果建模领域的进展,指出因果结构模型,由于其清晰的因果语义学和强大的因果图建模和转换能力,有望为先天/后天之争的解决提供一个有效的形式工具。第五节简要谈及人类对于演化过程的因果认知,指出因果结构模型可以为人类的因果认知中的规范性面向提供一种自然主义的诠释。此时,生物学中的因果张力才能被最终消解。
二、先天/后天之争与发育系统论
生物的性状是被先天决定的吗?关于这个问题,古希腊有预成论(Preformationism)与后生论(Epigenesis)之争。前者认为,生命个体预先存在确定的形式(form),发育仅仅是“小人”不断变大的过程。后者以亚里士多德为代表,认为生物的形式在发育过程中才逐步实现。但亚氏认为,父本的遗传物质提供了某种形成原则(form-giving principle),决定了个体的本质形式。这里,形成原则取代确定的形式,仍然预先决定了个体的发育和性状。到1900年,孟德尔的工作被重新发现后,经典遗传学用基因概念来指称引起表型的遗传单位。1953年,脱氧核糖核酸(以下简称DNA)分子结构的发现把基因锚定到了具体物理结构上。自此,现代遗传学基本成型,DNA序列精确编码了关于发育的信息,其分子结构保证了遗传信息的忠实传递。再一次,基因取代形成原则,预先决定了发育及其结果。这是现代版的预成论,有时也被称为基因决定论。
然而,当我们把目光投向有机体的整个生命周期时会发现,发育过程并非由DNA分子单独决定;后天因素,如体内的蛋白质、RNA等以及体外的温度、湿度等环境条件,也是导致表型性状的原因。2000年,当人类基因组计划初步完成之时,科学家便意识到,完整的基因序列并不能揭示人体的奥秘,更重要的工作是理清DNA与其他生物分子之间如何相互作用。几乎在同时,以科学哲学家苏珊·欧亚马(S. Oyama)为首的“发育系统论”(developmental systems theory)试图从根本上瓦解基因决定论。他们指出,基因只是发育的原因之一,非基因因素同样为发育提供了资源,两者的因果地位是等同的(“等同论题”,parity thesis)。等同论题可以用乳糖操纵子的例子来阐明(见图1)。
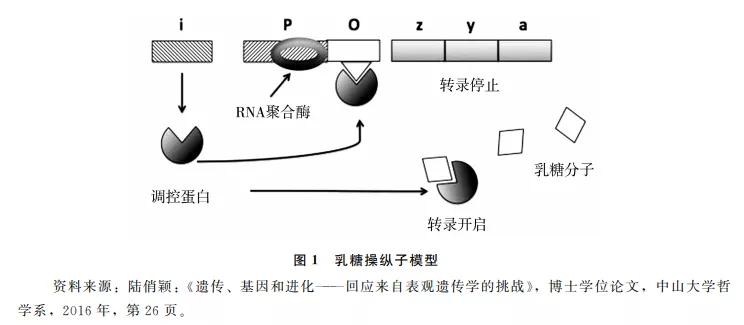
微生物学家对大肠杆菌的研究发现,存在一些DNA序列可以对编码蛋白质的DNA序列进行调控,从而抑制或激活后者的表达。如图1所示,上方的长序列代表一段DNA分子,其中z,y,a为结构基因,编码了分解乳糖的酶;P为启动子,结合RNA聚合酶后可以启动下游基因的转录;i为调控基因,其编码产生的调控蛋白可以结合到操纵子O上,使转录停止。当环境中乳糖含量较高时,调控蛋白多与乳糖分子相结合,使之较少结合到操纵子上,下游的转录被启动,产生了分解乳糖的酶,最终使乳糖含量维持在较低水平。发现者因此认为,基因不仅包含了编码蛋白质的蓝图,还可以调控结构基因的表达。类比于计算机,整个基因组可以根据不同的输入(乳糖浓度的高低)给出不同的输出(调控蛋白是否结合操纵子以终止转录),因此可以看作是一个集蛋白质合成与调控的“遗传程序”。
在发育系统论看来,上述对于乳糖操纵子模型的因果解读存在问题。乳糖调控系统是一个复杂的因果反应网络,截取其中的不同节点,可以看到不同因素的作用:若把乳糖浓度等环境因素作为背景条件,可以看到基因的重要影响;若把基因作为背景,则会看到环境对于结果的影响。乳糖操纵子模型的发现者选择了后者,得出了基因决定论的结论。但我们原则上也可以选择前者,以强调环境的决定性作用。更一般地说,为基因的因果重要性辩护的任何论证,同样适用于环境,因此没有理由认为基因比环境更具因果优先性。如果这个结论成立,那么对于基因与环境、先天与后天(nature/nurture)的二分也就失去了基础。此时,发育更像是一场爵士乐演出, 演奏者根据已有的旋律,即兴创造出相应的节奏和音符,构建起一个又一个的新主题。在表演结束之前,没有人知道整个音乐将以何种形式呈现。在这个意义上,先天与后天的界限被打破,发育成为一个真正的“后生”过程。
发育系统论的等同论题是有道理的。然而正如上一节所述,发育视角关心生物的结构如何产生,但这并不构成我们有关个体发育的全部理解。毕竟,在当代生物学中,“除非从演化的视角看,否则生物学的一切都无法被理解”。正如演化生物学家梅纳德·史密斯(J. M. Smith)指出,“一个DNA分子具有特定的序列是因为它决定了特定的蛋白质,但是一朵云是黑色的并非因为它能预测下雨”。仍然以类心脏的演化为例,类心脏结构的产生至少部分地由基因决定。假设给定正常的环境条件,拥有基因G会使个体产生类心脏。自然选择对于泵血功能的青睐使拥有基因G的个体被保留下来,基因G作为个体的一部分也被保留下来。正是因为基因G能够表达类心脏,所以它现在存在。在这个意义上,拥有基因G的个体“理应”表达类心脏。这使被选择的基因,与环境相比,拥有了一种因果特殊性:基因携带了由自然选择所赋予的内在目的,这个目的“预先决定了”发育的结果。
可以看到,一方面,根据发育生物学和分子生物学,基因和环境都是发育的原因,基因本身并不能预先决定发育。另一方面,演化历史对于生物功能的选择赋予了对应的遗传物质以特定的内在目的,奠定了基因决定论的直觉基础。这一看似的矛盾揭示了遗传物质——基因——在发育视角(结构导向)和演化视角(功能导向)下的因果张力。因此,先天/后天之争的彻底解决还要依赖于近因与远因之交互作用的澄清。事实上,这一探索在遗传分析的方法论讨论中已然开始。
三、遗传分析与差异制造者
先天/后天之争在各个分支领域,如行为遗传学和心理学,有更为具体的讨论。这些讨论主要聚焦于遗传分析的方法及其因果诠释。遗传分析的基本想法是,把群体中导致表型差异的遗传原因和环境原因区分开来。通过方差分析的统计学方法,可以估算表型差异中由遗传差异所贡献的占比,即遗传率(Heritability)。一般认为,遗传率衡量了表型在多大程度上由先天因素决定。这种因果诠释遭到了许多批评。早期的著名案例发生于20世纪70年代。亚瑟·詹森(A. Jensen)通过方差分析得到了某群体智力的遗传率为80%,因此声称“大约80%的智商差异具有遗传基础”。此观点受到了理查德·列文廷(R. Lewontin)的强烈批评。后者指出,遗传率所能推断的因果结论极为有限,并不能得出詹森的结论。列文廷之后的工作重在强调,发育是基因和环境互动的结果,他也是之后“交互论”(interactionism)的代表人物。


首先,激进的交互论者指出,表型是基因和环境相互作用的产物,两个原因原则上不能被分开来对待,公式(1)中对于基因型方差和环境方差的分解没有合理的基础。这个批评可以被轻松回应。简单来说,我们可以在承认基因和环境都是表型的必要原因的基础上,询问哪个原因更为重要。公式(1)实际上预设了两者的必要性。即使群体中表型的差异仅仅由其中一个原因的差异引起,另一个变量的存在仍然是必要的。这一点也由列文廷明确提出。列文廷区分了两类因果诠释。一类将基因和环境视为“相互排斥的替代原因”,并询问是哪一个导致了表型;第二类则认识到两者都是必要原因,并询问各自的相对重要性。遗传分析所针对的是第二类诠释方式。
温和的交互论者指出,遗传分析根据群体的数据得到因果的结论,这使结论无法应用于任何单一的个体。即便我们通过遗传分析得到了一个较高的遗传率,由于不涉及基因和环境的相互作用机制,所以不能得出任意个体的表型主要由遗传决定的结论。对于关心发育的生物学家来说,表型在多大程度上由基因决定的结论,并不能对真正的发育过程给出任何有效的因果信息。相反,他们真正想要的是,发育系统的各组成部分产生某种表型的具体机制。尽管存在这个批评,遗传分析在数量遗传学、行为遗传学和流行病学等生物统计学领域仍然有着广泛的应用。这场争论似乎由领域的分工被自动解决了。詹姆斯·泰伯里 (J. Tabery)基于这一事实,区分了群体层面的演化解释框架和个体层面的发育解释框架。
例如,当生物学家问,为什么有些人会出现抑郁症?这个问题可以有两种解读:
问题(1):抑郁症如何通过生物化学途径产生?
问题(2):为何有些人,而不是其他人,患有抑郁症?
泰伯里指出,交互论者关心的是问题(1),即特定基因型与特定环境的相互作用如何导致了抑郁症。要回答这个问题,任何与抑郁症相关的因素都要被纳入考虑范围,以刻画出确切的发生机制。泰伯里称此方法论为“机制阐明法”(mechanism-elucidation approach)。另一方面,演化论者关注的是问题(2),即造成种群中个体差异的原因是什么。此时,只有真正导致了表型差异的原因才会被纳入考虑。泰伯里称之为“差异划分法”(variation-partitioning approach),这正是遗传分析的方法论。两类问题对应两种方法论,遗传分析无法回答问题(1)并不构成对于其本身的批评。
上述两类问题正好对应迈尔的近因(如何问题)和远因(为何问题)。肯尼斯·沃特世 (K. Waters)对两类问题所针对的因果概念给出了进一步的分析。沃特世的工作从伍德沃德的干涉主义因果理论作为起点。干涉主义强调反事实干涉概念在因果探索中的重要性:面对复杂的自然现象,科学家通常会固定其他条件,干涉某个感兴趣的变量,如果干涉导致了另一个变量的变化,那么前者便是后者的原因。沃特世认同干涉主义将原因作为差异制造者(difference maker)的想法,但他认为,干涉主义不能解释一类特殊的问题。这里,沃特世引用了著名的火柴案例。玛丽在黑暗的房间中点燃了一根火柴,我们问:火柴燃烧的原因是什么?根据伍德沃德的理论,(其他条件不变)假如玛丽没有划火柴,火柴就不会燃烧;假如房间中不存在氧气,火柴就不会燃烧。因此,“氧气的存在”和“玛丽划火柴”都是“火柴燃烧”的原因。然而,在日常生活中,我们通常的回答是:因为玛丽划了火柴。
沃特世指出,为了解释上述日常直觉,我们需要区分两类原因:潜在的原因和实际上起作用的原因。某时刻玛丽没有划火柴,下一时刻玛丽划了火柴;对比这两个情境,玛丽划火柴的动作是火柴被点燃的实际原因。换言之,“氧气的存在”和“玛丽划火柴”都是火柴燃烧的潜在原因,但只有后者才是那个实际上使结果不同的原因。沃特世将潜在原因称为“潜在差异制造者”,实际原因为“实际差异制造者”。前者要求满足伍德沃德的反事实条件,后者要求实际上带来了结果的变化。沃特世继而指出,对于交互论者如分子生物学家和发育学家来说,他们想要确认所有可能影响发育的因素,即“潜在差异制造者”,并且理清各种因素在不同时间不同空间的相互作用模式。对于演化论者来说,他们关心的是种群中不同表型的占比如何通过自然选择作用等发生变化,即找到“实际差异制造者”,并分析其因果影响的大小,这里的主流方法便是遗传分析法。
沃特世的分析基本符合目前学界的共识,即方差分析可以提供性状演化的“实际差异制造者”(即远因)的信息,但无法提供关于性状发育的“潜在差异制造者”(即近因)的解释。然而,两种原因的概念区分并没有回答两者的内在联系。“潜在差异制造者”包括了“实际差异制造者”,但沃特世没有回答:哪些“潜在差异制造者”在什么情况下会成为“实际差异制造者”?我们仍然可以问:近因与远因的交互点究竟在哪里?更一般地说,基因对于表型在演化与发育中的因果张力并未消解。
四、因果张力与结构模型
先天/后天之争在具体方法论上的争议,使讨论从抽象的理论争论转变为了对因果概念更为详细和系统的辨析。学界逐渐意识到,单纯的概念区分并不能彻底解决一系列因果相关的问题,对于基因因果性的形式化研究是亟待完成的工作。在因果形式化议题上,生物学哲学近期主要有两大进路:一是运用数学工具梳理基因的因果特点,二是构建演化和发育的整合因果模型,而因果结构模型可以为这两条进路提供一个基础的形式工具。
第一条进路的奠基工作由伍德沃德做出,他基于干涉主义的因果理论归纳了生物学中尤为重要的三个因果特征,即稳定性、相称性和特异性,并给出了较为严格的概念定义。在此基础上,其他学者展开了针对这些因果特征的形式化工作。例如,保罗·格里弗斯(P. Griffiths)等人运用信息论工具构建了衡量因果特异性的数学方法。在正常情况下,基因的因果特异性要远远高于环境的因果特异性,这便部分地解释了基因的因果特殊地位。然而,这些工作所运用的数学工具(如信息论、概率论等)本身没有对应的因果语义,加之不同的学者会选择不同的数学工具。因此,一套完整的逻辑语言及其因果语义诠释的缺失,导致这些分散的形式化工作无法被系统地整合起来。
一个可行的思路是,首先给出与伍德沃德的干涉主义相对应的形式化系统,在此基础上,发展出不同因果特征的形式定义。幸运的是,在因果建模领域,作为干涉主义的一种形式版本,因果结构模型的工作重塑了我们对因果性以及统计与因果的认知。其当代的发展主要由卡耐基梅隆大学的克拉克·格莱摩(C. Glymour)团队和朱迪亚·珀尔 (J. Pearl)做出。以后者的工作为例,珀尔用“do-算子”定义了形式化的“干涉”概念,据此建构起了关于因果的形式语义学,给出了严格的基础概念,如因果关系、因果力的度量等,使因果推理的形式化计算成为可能。巧合的是,珀尔近期运用了因果模型来解释火柴案例中的因果直觉。基于他的因果形式系统,珀尔得到了衡量“因果充分性”的公式,即充分性概率(probability of sufficiency,PS)。在火柴案例中分别计算出“氧气的存在”和“玛丽划火柴”对于“火柴燃烧”的PS值。其结果显示,“玛丽划火柴”的PS值要远远大于“氧气的存在”的PS值,即“玛丽划火柴”是“火柴燃烧”的更为充分的原因。
相比于沃特世的概念区分,珀尔的工作用PS值将两种因果概念放置于同一个度量中,呈现了“实际差异制造者”和“潜在差异制造者”的可能连接点。相比于格里弗斯等人的工作,珀尔的工作有一套完整的形式语义系统作为支撑,在理论辩护上更具优势。另外,珀尔得到的结果作为整体语义系统的一部分,还可以连接其他的因果度量,因此具有更广泛的应用范围。可以预见,因果模型的运用有望为伍德沃德所总结的生物学中的不同因果特征给出一个整体性刻画,也许还会修正对于因果特征的理解,并提示一些未曾想到的细节问题。
关于生物因果性的第二条进路,试图从根源上解决基因的因果张力问题。尽管人们普遍接受泰伯里和沃特世的结论,即生物演化论(演化问题)和分子生物学(发育问题)具有两种不同的解释框架和方法论。但若能构建出发育与演化的整合模型,则有望消解基因在两个维度的因果张力。这条进路最初由日本学者大冢俊(J.Otsuka)尝试,他以演化论中的普莱斯公式(Price Equation)作为研究起点,运用因果图的方法构建了发育和演化的整合模型,试图揭示近因与远因的交互影响。但是因果语义系统的缺失,使之只能在普莱斯公式的基本思路上进行分析,即以适应度(fitness)为核心变量,尤其关注演化的维度,因而无法将影响性状的其他因素以等同的方式纳入进来。
因果图的思想最初来源于20世纪20年代生物统计学家休厄尔·赖特(S. Wright)的路径分析(path analysis)。在当时,这是运用统计学解决因果问题的创新尝试。赖特的基本想法是,用带箭头的连线表示变量之间的因果路径,并根据观测或实验数据估计每条路径的因果力,即路径系数。由此得到的“路径图”便是因果图的前身。路径分析在统计学之后的发展中被暂时遗忘,取而代之的是针对“相关性”的方差分析、回归等技术。但有趣的是,赖特对因果问题的思考方式在当代产生了持久的影响,有望为方差分析方法弥补因果诠释上的缺憾。其最为重要的影响便是因果结构模型。珀尔将因果图的思想整合到了因果形式系统中,造就了其强大的因果建模和因果计算能力。
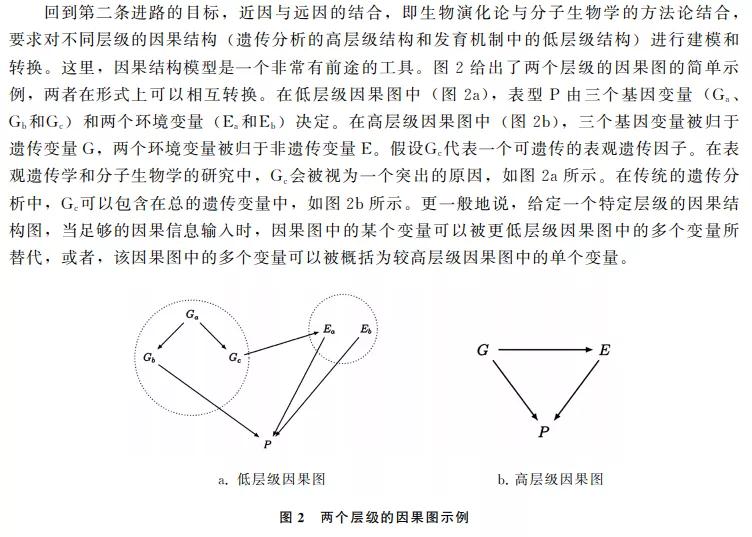
当然,有效的建模工具需要有效数据的输入,生物学的新近发展提供了建模所需的大量数据。一方面,像GWAS(全基因组关联研究)这样的新技术被发展起来,可以精确定位影响性状表型的大量DNA序列,即定位传统遗传分析所追寻的“实际差异制造者”。另一方面,分子生物学的知识积累为阐明表型在发育过程中产生的机制提供了大量有用的信息,即定位“潜在差异制造者”。这两方面的信息整合,恰好能为不同层级的因果图建模提供数据来源。因果结构模型所具备的因果图的跨层级转换能力,可进一步促进演化论与分子生物学的学科交叉与互动,并有望为基因的因果张力的消解提供一个有效的形式工具。
五、生命目的与因果认知
达尔文的自然选择为生命的目的提供了机械论解释,是当代生物学最为突出的因果特征。然而,这一特征却与发育的交互论产生了看似的冲突。从演化视角来看,由于现存的某些基因具有一个基于自然选择的演化历史,该历史赋予了它们指向特定表型的目的性。换言之,在生命孕育之初,基因便“预先决定”所对应的表型,不管事实上这个表型是否被实现。这种关于基因决定论的理解也被称为“目的论语义”(teleosemantic)进路。该进路基于一种与语义特征的类比论证,例如,两者都具有意向性和可错性特征:语义内容总是“关于”某些目标对象,若命题未能成功指向目标对象,则命题为假;被选择的基因总是“关于”特定的适应性性状,若未能实现该性状,则演化历史所赋予的目标就没有被达成。
这种类比论证当然促进了我们关于演化视角下基因对于表型的因果理解,但是我们仍然可以追问:自然选择究竟在何种意义上赋予了基因以生命的目的?为了回答这个问题,我们还是需要回到基因的因果角色中。如果我们承认,演化的过程本质上是一种物理的因果过程,那么基因实际上不可能真的携带任何的目的。基因决定论中有关生命之目的的直觉只能来自人类对于因果过程的认知。笔者认为,只有把视角转向关于我们对于基因在演化和发育维度的因果认知时,关于基因的因果张力才能被最终消解。
近年来,关于因果认知的研究越来越与因果结构模型相关。一个公认的事实是,当某个结果由多个原因导致时,人类倾向于对不同原因的因果力进行排序,就像把“因果分数”归于每个原因。因果建模领域多用概率来衡量因果力,上一节所举的珀尔的“充分性概率”(PS)便是一个典型例子。从因果认知上看,一般认为,我们对因果力的判断近似于我们对未来“干涉”的期望。因此珀尔的PS也能用来衡量我们对干涉的结果的预期。在火柴案例中,由于“玛丽划火柴”是“火柴燃烧”更为充分的原因,即“玛丽划火柴”充分导致“火柴燃烧”的概率较高。我们会期待,在未来,如果玛丽再次划火柴,那么火柴燃烧的概率会很高。假设这个概率接近最大值(1),那么我们会认为,“玛丽划火柴”理应导致“火柴燃烧”;如果结果火柴没有燃烧,一定是哪里出了问题。这便为因果认知中的规范性面向提供了形式化的解释。
相应地,从演化维度来看基因对于表型的因果力:如果某基因在自然选择之后,其对适应性表型的因果力大大提升,并且几乎达到了最大值(1)。那么我们会对未来的“干涉”结果设定一个“规范”的预期,即该基因在未来“理应”表达适应性表型;如果事实上该基因没有表达适应性表型,那么我们会认为,自然选择所赋予的目标失败了。在这个意义上,人类对于基因在自然选择中的因果认知,产生了被选择的基因对于适应性表型的规范性认知。只有在此时,我们才能为生命之目的提供一种彻底的自然主义诠释。
(本文注释内容略)
原文责任编辑:王志强
扫码在手机上查看












